Chapter 5: Corrections on Evolution
Chapter Summary Scientist get very little right about early evolution, missing anything that cannot be studied in a laboratory; and they don't study broad enough areas to understand the complex interactions. They assume an asteroid destroyed the dinosaurs, but it was grass which overcame oppressive brush of nonwoody plants eliminating dinosaurs and shaping modern biology. Even if an asteroid or volcano speeded up the extinction, it was grass which shaped the result and would have produced the same result eventually. The nature of the transformation shows the influence of grass in promoting modern biology while eliminating vast amounts of primitive biology. ◆ ◆ ◆ ◆ ◆ ◆ ◆ ◆ ◆ ◆ ◆  Soil Formation Scientists are mystified by the Cambrian explosion of life. About 543 million years ago, the number of species vastly expanded. The most likely explanation seems to be that a planet exploded between Mars and Jupiter where the asteroid belt exists, and good quality clay was layered onto the surface of the earth. Numerous critical minerals were probably added to the earth’s surface also. Since the Cambrian explosion of life occurred first in the oceans, it was probably the deposition of minerals in the oceans caused by the planet exploding that created the initial result. There was probably very little for usable minerals available to biology prior to that time. There are two reasons why scientists cannot draw these conclusions. One, they don’t assume that a planet exploded, at least at this time. And two, they assume soil was produced by plant roots breaking down rocks. This means there was no soil until long after plants with roots evolved, which was around 450 million years ago. The concept is totally ridiculous, as plants with roots cannot grow on rocks; they need soil. Apparently, shale was the closest thing to soil to be formed while the earth was created. Oceans could erode the shale creating soil-like sediments. This material probably did not get to the land surface until recent times, because major upheavals of continental plates would be required. Of course volcanoes contribute something to soil, but these minerals need improving through biological activity and mixing with other soil. More significantly, there were no volcanoes until about 300 million years ago, because the tectonic plates were extremely thin. They keep getting thicker as the planet cools. Volcanoes cannot be produced until the tectonic plates are thick enough to get damaged as they bump into each other, basically by sliding over and under each other. Since tectonic plates were very thin prior to 300 million years ago, there would have been nothing for mountains, and the terrain would have been close to flat.
Conifer trees evolved about when tectonic activity began creating volcanoes, 300 million years ago. The driving force for conifers was apparently the formation of hills, which allowed plants to separate into nonwoody types in the low areas and woody conifers on the hills. Dinosaurs ate nonwoody plants only. Biologists cannot produce a realistic explanation for the size of dinosaurs. There is always environmental pressure creating significant evolution. The obvious force for dinosaur size was that walking through brush is extremely difficult. It takes a lot of power, which takes a lot of size. Nonwoody brush would have had no open space under a canopy, as modern forests do. So dinosaurs would have had no choice but to tromp through heavy brush. The brush would not only have provided food but would have also protected eggs and young dinosaurs from predators. The theory that dinosaur extinction was caused by an asteroid impact lacks too much. Even if such an asteroid were involved, it would have been a minor part of the process. Dinosaurs were due to be replaced, as evolution does not stand still. It constantly gets more specialized and efficient. Dinosaurs should not have lasted long with their extremely crude characteristics and cumbersome size. It was the nonwoody brush that locked dinosaurs and much of their ecosystem in place. The brush kept mammals small, as they had to move under and through the brush. All the while, the flowering plants were slowly and invisibly setting the trap for dinosaurs—not only the dinosaurs but the nonwoody brush as well. The white knight, the Robin Hood, saving biology from the scourge of nonwoody brush was grass. The most significant property of grass is that it absorbs water above the ground in addition to through the roots. This allows grass to thrive on much less precipitation than other plants. The long, pointed leaves allow dew drops to run down to the base, where moisture is absorbed. To outcompete other plants, grass formed an impenetrable mat on the surface. Very few nonwoody plants survived the evolution of grass, and dinosaurs went out with the nonwoody plants. Dinosaurs could not survive grass, because they had to eat nonwoody plants; and grass would have exposed their eggs and young to predators. If an asteroid impact was involved with the extinction of dinosaurs, it would have been a minor contribution to what would have happened anyway. The evolution of grass is visible in a newly discovered dinosaur (Anzu wyliei) which looks like a large chicken due to its thin, long legs. Long legs and light weight are needed for walking through grass. This dinosaur was one of the last to exist before they all became extinct. It means grass was significantly developed when dinosaurs died out. Grass shaped modern biology. Mammals could walk over it and through it, which freed them to diversify. Flowering plants found space around the grass and sometimes within it. They evolved into a new type of woody plants which became broadleaf trees. Flowering plants evolved sugary substances, which allowed yeasts to evolve from filamentous fungi. Fungi also evolved into modern mushrooms which could grow within the grass. All this began 65 million years ago. The official story is that the extinction of dinosaurs left "space" for mammals to evolve. How could space be relevant? A dinosaur takes up a few cubic feet of space, and the earths surface has quadzillions of cubic feet of space. Are we supposed to read something into space? Is that supposed to be science— reading something into a word? That's the point. Power mongers replaced science with word salad which goes on and on. Mammals were exploiting dinosaurs, as they would have been relying upon dinosaur eggs as a primary source of nutrients. After the dinosaurs were gone, most mammals had to adapt to eating grass. Predators could then follow the grass eating mammals. When significant terrestrial life began, shortly after the Cambrian Explosion in 543 BP, fish were soft bodied, as bones had not yet developed. When a planet exploded in the asteroid belt and left good quality soil on the surface of the earth, amphibians formed and moved onto land. They created internal bones. It took gravity on land to cause bones to form. Evolution biologists can't figure out why evolution created extreme diversity during the Cambrian Explosion. They can't seem to quite grasp that evolution does not create diversity. Evolution does not shape evolution. Environments shape evolution, and environmental change is required to create diversity. The extreme diversity during the Cambrian Explosion had to result from environmental change. How then did fish acquire bones? They would have gotten bones from amphibians which migrated back into the water. Then the boney fish migrated back out of the water onto land (400-300 million years ago) and evolved into mammals. This pattern is evidenced by recent studies which show the limbs of mammals evolved from fish fins (Science Daily, Jan 21, 2014). The amphibians which stayed on land evolved into reptiles. Reptiles are not as advanced as mammals, because they did not go through the drastic environmental changes of migrating back into the ocean and back onto land, as mammals did. The experience of going back into the oceans and back out created much more evolutionary change for mammals than reptiles experienced. This pattern of migration is indicated by the two dots which humans have below their front teeth. The dots can be felt with the tongue. The dots are remnants of hooks used by ancient fish, which means mammals evolved from boney fish. Going into the oceans and back out did not in itself shape the evolution of mammals. Instead, it made the DNA more prone to evolution, presumably by stripping proteins away from a large part of the DNA so it could evolve more freely. The amphibians which evolved into reptiles did not undergo as much change as mammals, as their DNA would have been less available to evolution. The proteins which protect DNA did not disappear from the mammalian line; they mysteriously acquired the ability to allow evolutionary change while continuing to resist undesirable mutations. Evolution does not occur as significantly or rapidly in the oceans as on land, because the oceans are extremely stable. It takes environmental change to produce evolutionary change. Tides create more change near the shoreline. For this reason, most of the diversity in the oceans resulted from land creatures continually adapting to the oceans throughout evolutionary history. Mammals are still adapting to the oceans and creating a wide variety of sea creatures. One of the most astounding products of evolution is the bacterium Pseudomonas fluorescens (P.f.). It never branched on the evolutionary tree; it just kept adding more functions. About a 800 million years ago it swam in an ancient sea using two polar flagella which rotate. Polar flagella would have evolved before terrestrial life began, 543 million years ago, because exotic structures were easy to produce during early evolution. Filamentous fungi show extremely frivolous structures which evolved early in evolution, such as innumerable variations of the clamp connection. Not only was there less competition and related demands early on, but less specialization allowed more varied alternatives. All biological life is differentiating itself out of existence through over-specialization, which is why so many species die out when environmentally stressed. Biological life could end in 10 million years or less due to over-specialization. There is another type of flagella called peritrichous flagella, which surrounds some bacteria. That flagella is designed for moving through viscous liquids, and it is more recent. The difference stems from modern biology creating viscous liquids with nutrients, while in the ancient world water with few nutrients was the primary liquid for biology. Scattered nutrients in an ancient sea gave P.f. a more versatile nutrition and physiology. Now days, bacteria specialize more, because they have specific habitats with consistent nutrients. Evidence of P.f. moving out of an ancient sea onto land is in the fact that it is the most common and well adapted soil bacterium. P.f. adapted to soil better than any other bacteria, because it was the first bacterium in the soil. Pseudomonads are linked together in pairs under nonaqueous conditions, which is the basis for the name. This allows one of the bacteria to break open and release enzymes near the other one. Whereas molds produce enzymes on their surface, bacteria have to break open to release enzymes. P.f. breaks down proteins in the soil, which is highly significant where ground freezes. Ice crystals break apart most cells in the soil including some plant roots, fungi and insects. P.f. is the first bacterium to act upon the resulting cell debris in the spring. But it will also grow on extremely diverse nutrients. The isolation medium for it is glycerol and nitrate. Nothing else noticeable grows on that combination. Under nonaqueous conditions, P.f. produces a pink, nonwater-soluble pigment. This pigment shows up when growing P.f. on glucose and agar. Most bacteria would die for a pigment, but pigments are usually too extravagant for them. P.f. has two. Pigments allow insects to see and land on bacteria; so they to pick up the bacteria and carry them around. Also indicative of evolutionary age is symbiosis. The oldest species accommodate their environment the best. P.f. is like a slave for plants feeding them organic nitrogen. For example, when grains such as wheat are planted in the spring, the leaves start to turn yellow due to a shortage of nitrogen, because P.f. hogs up the nitrogen early in the spring. After about a month of growth, the leaves turn dark green, as nitrogen becomes available. P.f. autolyzes (self-destructs) and releases nitrogen to the grain in the nick of time. Plant roots excrete acids from the TCA cycle, which increases bacterial growth around them and induces some autolysis by the bacteria to feed on them. Mushrooms can then exploit the nitrogen along with the plant roots. With autolysis providing complex nitrogen, a diverse population of bacteria develop around plant roots. Even morel mycelium is drawn to plant roots because of the bacteria, while its normal growth pattern is to spread over several meters of space in sandy soil feeding on P.f. The polar flagella of Pseudomonas fluorescens uses several proteins to create the rotary motion, and chemical energy as ATP is required. Respiration now uses similar rotating proteins to create ATP. This parallel structure would have once been called convergence of evolution. But it is transposed evolution. Respiration would have acquired the rotating proteins from the locomotion of Pseudomonas fluorescens. The transformation would have occurred within the cells of Pseudomonas. The bacterium has highly varied nutrition and physiology. The rotating proteins are not essential for ATP synthesis; they simply speed up the process by moving the substrate molecules into and out of the reaction site faster. Without the rotating proteins, diffusion would be required for moving the molecules around. Every metabolic reaction would benefit from rotating proteins moving the reactants in place, but the process is too elaborate for anything but respiration. Speed is important for respiration, so organisms can get more done faster without running out of energy. Such extreme evolution cannot occur where metabolic activity is highly active and complex. There are too many inter-dependent activities to allow such drastic changes. But there are no competing influences for the evolution of locomotion. So the rotating proteins could freely evolve without disruption while producing locomotion. Then adapting the rotating proteins to respiration would be simple enough that it could occur without disrupting essential metabolism. Pseudomonas could gain energy through other methods while the new form of respiration was evolving in its metabolic machinery. Respiration through simple diffusion would have existed before the rotating proteins were added. The upgrading of respiration would have occurred about 700 million years ago plus or minus 100 million years. The new form of respiration involving rotating proteins was so advantageous that it was carried into the cells of soft-bodied fish as bacterial parasites. These then evolved into mitochondria. Another example of transposed physiology is fat production. There was no fat as energy storage before modern biology began and dinosaurs died out. Storage molecules as starch or fat did not exist. All evolution was being held back by nonwoody brush. When dinosaurs died out, grass shoved out the nonwoody brush and freed up evolution for all species. As plants produced sugary substance, yeasts evolved from molds. Yeasts acquired a round shape, which improves absorption in liquids; and they gave up the most demanding characteristic of molds, which is the ability to tolerate dehydration while growing on surfaces. The primary feature of yeasts is that they repress TCA enzymes while glucose is available. This allows them to metabolize glucose rapidly preventing competitors from using it. The acetate resulting from glucose metabolism is channeled in three directions. Some is excreted as acetic acid and some as ethyl alcohol. These substances inhibit growth of bacteria and molds. Production of acetic acid and alcohol are phenotypic variations. Oxygen does not have to be absent to produce alcohol. But unlimited amounts of these substances cannot be produced, so acetate is also channeled into fat production. Fat is created from acetate molecules linked together. Yeasts often store up to 40% of their cell mass as fat. When glucose gets used up, yeasts induce formation of TCA enzymes and re-metabolize the acetic acid, alcohol and fat which they created earlier. Other species do not have repressible TCA systems. Their TCA systems are in place all of the time. The fossil evidence of yeasts only goes back 50 million years (1). This would be when plants produced sufficient sugar to promote yeast evolution. Fat would presumably have been nonexistent prior to that time. The bone structure of dinosaurs and early mammals indicates that these animals were not producing fat. They are too tall for fat production. Dinosaurs could have benefited from fat, because they needed a lot of weight to go through the thick brush. If they had fat, they would have been shorter and heavier like elephants. The early mammals were also longer legged than they would have been if they stored fat. Modern mice are much shorter, because they are heavier due to fat storage. It was the repressibility of the TCA system which allowed yeasts to evolve fat production. No other species would have been using repressible TCA systems, and therefore, it is most likely that no other species would have evolved fat production. After yeasts evolved fat production, the related genes would have been transferred to other species by viruses. If this path of evolution is correct, animals would not have had fat during the first several million years of modern biology beginning 65 million years ago. Evidence of the evolution of fat may be visible in the fossil records. It would show up as a shortening of leg length for mammals. Mammals should have gotten shorter and heavier-boned when they acquired fat production. Another line of evidence of the evolution of fat is in the cottonwood and willow trees. These trees stopped evolving significantly very early on, because they found a highly exploitable niche by drawing water from stable sources such as lakes or rivers. Their seeds are very small compared to modern seeds. Modern seeds store a lot of energy as oils. If fat production was possible when cottonwoods and willows evolved, they would probably have had larger seeds. Their small seeds indicate that fat production did not exist when they evolved. Cottonwoods and willows are more primitive than other trees in that they do not measure stress forces. Modern trees measure stress and widen where stress forces are greatest. As a result, all locations on the trees have an equal probability of breaking. Ancient trees including conifers do not measure stress. Their branches do not taper significantly beyond the effects of aging. Aging produces less taper than the measurement of stress. So conifers and cottonwoods have long branches without much taper. The willow adapted high flexibility to cope with stress. But the trunk of the willow is not flexible. As a result, willows are often found collapsed onto the ground and still growing. In other words, it is the primitive nature of these trees which resulted in small seeds, not some unseen advantage.
There are always environmental forces creating evolutionary change. What happened at that time is that tectonic plates were getting thick enough to create significantly sized hills as they bumped into each other. Rain water running off the hills discouraged the growth the nonwoody brush and left open space for new plants to evolve. Conifers filled that space on the tops and sides of hills, while nonwoody brush stayed in the lowlands. The conifers would have created a favorable environment for mushrooms. Plants were producing a lot of round aggregates of cells back then and appeared to be evolving into fungi. (See "The Biology and Evolution of Fossil Plants", 1993.Thomas N. Taylor and Edith L. Taylor.) My impression is that numerous different fungi evolved from plants independently after terrestrial life began. So why do they have similar cell walls if independent? This question would have been unanswerable a couple decades ago, but what scientists have learned from genetic engineering is that horizontal transfer exchanges genes between all species. Viruses, bacteria and fungi serve as the carriers. This is noticeable in wildflowers. They all seem to have a combination of the same basic scents, even when they are not related. Horizontal transfer apparently carried some of the same genes throughout the wildflowers. This means that plants could evolve into a myriad of fungi by picking up the cell wall genes from other fungi. By evolving from plants instead of other fungi, more versatility was possible. Evolution cannot go backward from more specialization to less specialization. Restarting evolution from plants would be like starting from stem cells allowing greater diversity than all fungi evolving from one point. The puffball is extremely hardy. It likes to emerge from car trails on dry prairies. It can determine where trails are by sensing carbon dioxide. Grass is shorter there allowing spores to disseminate easier. The ground gets dry and hard very rapidly after rains, because capillary action is high in fine textured soil. So the mycelium grows for awhile during rains, and then becomes almost spore-like between rains.
These varied puffballs all appear to be phenotypes of the same genotype, because they appeared near each other within a few months. They would all be growing from the same mycelium, because puffball mycelium is so widely scattered that two sources almost never come together. It appears that the puffball can change its phenotypes without going through genetic recombination, unlike other species. It's possible that the mycelium sectors as a method of creating phenotypes. This would be like cells changing phenotypes to create tissues but then going beyond the tissue to create individual phenotypes. The puffball produces no visible gene exchange. In general, ancient species do not need much gene exchange, because they acquired very high survivability through versatility. Highly stable phenotypes appear to be a major element of the versatility of the puffball. It's lack of gene exchange creates a need for a few stable phenotypes which presumably always appear together. Puffball mycelium stays in one location for several decades, slowly expanding under the ground each year without ascocarps emerging for decades. The mycelium can travel a hundred meters along a car trail before puffballs emerge, and then they come up all along the mycelial trail. Not emerging while the mycelium expands appears to be a means of preventing predators or diseases from developing against the species. Such extremely ancient organisms produce that sort of versatility. The new locations are widely scattered and random. My guess is that the spores are pressure sensitive, so that only those that get high in the atmosphere can germinate. Perhaps other airborne mushroom spores require atmospheric conditions, of which low pressure is the only apparent mechanism, for evading diseases. Agaricus avoids growing in the same area from year to year. What keeps the spores from germinating in the vicinity of the mushroom which produces them? It might be atmospheric conditions being required to precondition spores for germination. Coprinus comatus, however, liquefies to drop some of its spores in the same area, which means its spores do not need to be exposed to atmospheric conditions. The advantage of reducing gene exchange is that mycelium patches can be widely scattered to prevent insects and disease from developing against the species. Without much gene exchange, a few highly specialized phenotypes can take care of the need for variation. The puffball is so advanced that it gets by with almost no gene exchange and only four phenotypes all springing from the same mycelial patch. By contrast, the morel produces unlimited phenotypes by randomly remixing a large number of characteristics creating nonfunctionality for most of the resulting phenotypes. Each spore pair of the morel is a different phenotype with little probability of any two being the same. There are, of course, a wide variety of genotypic variants evolving for the puffball in response to the highly varied ecology that modern biology created. Throughout biology, there is a central line of evolution for types and a constant splitting off into branch types, most of which die out rapidly. With diseases, the branch types tend to be more deadly, while they die out rapidly, as small pox did. Long term survival requires a degree of compatibility with other species, which tends to promote symbiosis. Also ancient is the mushroom Boletus edulis (B.e.), which is highly valued by chefs. B.e. does not have gills. Instead it has tubes under the cap with spores in them. When B.e. evolved, mammals were small and shrew-like. Nonwoody plants produced that result. These plants would have created a very thick brush. Dinosaurs had to be large to walk through that brush. Mammals had to be small to go under it. Mushrooms could not grow in that brush. The surface of the ground under the brush would have had a thick layer of undecayed debris. Puffballs probably grew around the edges of the brush feeding on bacteria associated with plant roots, while B.e. adapted to the roots of conifers. Wind cannot easily get into conifer forests, so B.e. could not exploit wind very well for spore dissemination. One reason why B.e. did not evolve better spore dissemination is because it must be perennial to grow in tree roots, so it is stuck in a fixed location from year to year. Since B.e. evolved in response to conifer forests, it probably has the same evolutionary age as conifers, which is 300 million years. It's possible that puffballs evolved on low hills before conifers, since they are not dependent upon tree roots. This would make the evolutionary age of puffballs more than 300 million years, but there is no evidence of exactly when puffballs evolved other than having ancient characteristics similar to Boletus edulis including tissue which does not easily dry out and evolving away flavor from young tissue, so animals do not eat it. Almost all terrestrial biology changed drastically as the dinosaurs died out. The drastic changes were caused by grass. Grass shoved out most of the nonwoody plants freeing both plants and animals from the oppressive brush that hindered growth for most species.
Some varients of Boletus edulis appears to not have changed significantly as modern biology replaced the primitive biology of the dinosaur years. B.e. has a bulb at the base of the stem, which does nothing to strengthen the stem, as it curves inward at the bottom. It's function would be to allow small shrew-like mammals to stand on the bulb while eating spores under the cap. Mice still eat the spores allowing the bulb to persist without evolving away. The spore area of B.e. has more flavor than the rest of the mushroom, and Europeans sometimes use the spore area separately for cooking.
When spore dissemination is limited, as with the bolete or the morel, changes in genotype tend to occur in local areas. But when spore dissemination is very good, as with the puffball, minor differences in genotype cannot develop, because one genotype will prevail over the others. Then phenotypic differences get more exaggerated, which appears to be the case with the puffball. As Boletus edulis gets large, the bulb on the stem disappears. This is because larger animals are then depended upon, and not a single molecule is wasted on a form that is not needed with such an ancient mushroom. Structural tissue is the same, and very unusual, for Boletus edulis and puffballs. The tissue has a gummy texture without fibrous material. This means water and nutrients must diffuse from cell to cell, which is a slow process. Therefore, it takes two weeks for the Boletus edulis sporocarp to form, but only one or two days for Agaricus. Bolete tissue is very slow at drying. As a result, boletes often survive for two months, while Agaricus tends to dehydrate and disintegrate in a few days. Both the bolete and the puffball evolved away all flavor from the structural tissue, so animals will not eat it. The flavor of Boletus edulis increases as the mushroom dries, and the structural tissue picks up flavor upon drying. This effect probably evolved to enhance the tendency of animals to eat the mushroom as it ages. When Boletus edulis is dried slowly it acquires more flavor than when it is dried rapidly, which indicates that a biochemical process is involved in enhancing the flavor. Gilled mushrooms cannot evolve away flavor, because there are too many molecules which would need to change, and evolution progressed to more specialized functions. Therefore, gilled mushrooms have to produce a toxin, if they need to keep animals from eating them. Pseudomonas fluorescens (P.f.) is 600-700 million years old in its recognizable features. Evolutionary age is not highly definable, since all species branch infinitely all the way back to pre-life chemicals. But a concept of age exists based upon recognizable characteristics that go back in evolution. P.f. has recognizable characteristics that go back that far. Its polar flagella would be that old. Before that, the bacterium would have existed in some form for at least a billion years. The age of the polar flagella of P.f. can be determined from the evolution of modern respiration. There are a group of rotating proteins that speed up respiration allowing animals to move faster than they could with simple diffusion supplying the energy. Those rotating proteins evolved from the polar flagella of P.f. This relationship can be stated as an unquestionable fact of evolution due to the similarities and physiology. One state must evolve from the other state, no options about it. The reason is because polar flagella were free to evolve without interference; while respiration proteins could not have evolved on their own due to demanding physiology. The physiology of respiration is locked into cell structure and requirements for energy which left no room for drastic alternatives. On top of that, the new form of respiration did not work its way into the old form of respiration. Instead it entered the cell as whole bacteria which transformed into mitochondria. This addition to cell structure allowed the new form of respiration to be added to the old form while maintaining a variety of methods of producing energy as ATP. Since P.f. has a history of at least a billion years as a bacterium, the evidence of its age is unmistakable. As P.f. adapted to various environments, it branched off into all of the other gram negative bacteria while leaving the original state as the main stem of gram negative evolution. All of the diversity of gram negative bacteria traces back to P.f., which probably includes Escherichia coli (E. coli) which entered the intestines of animals from their beginning, which would be about 500 million years ago. Since P.f. is much older than E. coli, E. coli would have evolved from it. Yet there is the claim by some persons that the gram positive bacteria are older than the gram negatives. Those persons do not give a credible explanation. It is widely known that P.f. goes back in some form at least a billion years. To get gram positives back farther would be assuming they include cyanobacteria, which is stretching the concept of gram positive bacteria too far. The concept of gram positive bacteria became highly developed while cyanobacteria were being called blue-green-algae. The modern form of gram positive bacteria (apart from Streptomycetes) clearly evolved with modern biology beginning 65 million years ago with the die-out of dinosaurs. Two major indicators are the simplicity of their characteristics and their affinity for carbohydrates. Biologists are unfamiliar with the transition that occurred 65 million years ago. They don't know that there were no storage molecules before then including starch, fats or free sugars. Circumstantially, there could have been traces of such evolution beginning in obscure locations such as flowers of rare plants, but this physiology was not spreading to other species to influence other organisms. When flowering plants began to produce sugary substances after the transition, 65 million years ago, the result was the evolution of yeasts and modern gram positive bacteria, as they adapted to the sugary solutions. Modern gram positive bacteria have an affinity for carbohydrates, while gram negatives have an affinity for nitrogen containing molecules. The reason is very obvious. Before the transition 65 million years ago, gram negative bacteria had to destroy other cells for nutrients, and they are designed to do that. Their cell walls chew through other cells upon contact, which makes them the dread of biology. Modern gram positives do not do that. They feed upon readily available nutrients of the carbohydrate type which are released by dead vegetation. Some of them later became pathogens, which might not look like a vegetarian diet, but other similarities show the same origins. Microbiologists are not aware of this difference in nutrition—affinity for carbohydrate vs. nitrogen containing molecules. Here is a major source of information which has been out of view of microbiologists: Many plants and most mushrooms protect themselves from the scourge of gram negative bacteria by causing gram positive bacteria to form on their surface. Excretion of carbohydrate would be the method of promoting gram positives on the surface. A highly visible example is green beans. After sitting around for awhile, they get coated with a buttery substance on their surface. It's edible and doesn't have much flavor. It would be a coating of gram positive bacteria, probably Bacillus cereus, though I have not tested it to find out what species it is. Microbiologists do not know what the evolutionary habitats are for B. cereus and B. subtilis, but circumstantially, those similar bacteria would have evolved on the surface of plants being promoted as protection from gram negative bacteria, while variants would have evolved on decaying vegetable matter. A highly informative bit of evidence is the morel mushroom. It has no such protection having evolved from a single-celled yeast a few months ago. As the morel ages, gram negative bacteria invade the tissue. Persons who eat old and deteriorating morels get sick from them, as the invading gram negative bacteria have an endotoxin in the cells walls. That type of toxicity never occurs with other mushrooms, as they protect themselves from gram negative bacteria, though some of them produce their own toxin for protection against predators. There are some very complex gram positive bacteria, such as Listeria, which appear to go back in evolution farther than the other gram positives. Listeria would have evolved from Streptomycetes, since it has a soil-adapted stage. Streptomycetes are filamentous bacteria which are highly specialized in breaking down cellulose in soil. They have been breaking down cellulose in the soil for 450 million years. They would have been the evolutionary source of modern gram positive bacteria which evolved on carbohydrates after the biological transition 65 million years ago. Streptomycetes would have evolved from fungi, which have cell walls similar to gram positive bacteria. The modern gram positive bacteria would then have evolved from Streptomycetes when modern biology began. A major element of the biological transition 65 million years ago was the development of flowering plants and sugary substances that they produced. Both yeasts and gram positive bacteria evolved on the sugary substances. The gram positive bacteria also fed on the yeasts, while the yeast protected themselves by excreting acetic acid and ethyl alcohol. The yeasts sort of won, but the gram positive bacteria diversified and adapted to other plant material. These changes modernized biology in very dramatic ways. Not only did mammals rise up to dominate the animal world and flowering plants emerge from obscurity to dominate the plant world, the gram positive bacteria rose up out of the soil as Streptomycetes and moved into the plant and animal world as modern gram positive bacteria, first as scavengers and then as pathogens. --------------- 1. Thomas N. Taylor and Edith L. Taylor. The Biology and Evolution of Fossil Plants. 1993. Prentice Hall, Englewood Cliffs, New Jersey. |
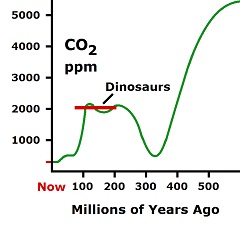 The beginning of volcanoes 300 million years ago is indicated by the CO2 graph which shows a restoration in the CO2 level at that time. Oceans continually absorb carbon dioxide and tie it up as calcium carbonate and limestone. Almost all CO2 was removed from the air 330 million years ago. In the nick of time, volcanoes added back significant CO2. Now the CO2 level is again dangerously low, as oceans continue to tie it up, while volcanic activity is decreasing due to extremely thick tectonic plates which do not easily allow lava to get through.
The beginning of volcanoes 300 million years ago is indicated by the CO2 graph which shows a restoration in the CO2 level at that time. Oceans continually absorb carbon dioxide and tie it up as calcium carbonate and limestone. Almost all CO2 was removed from the air 330 million years ago. In the nick of time, volcanoes added back significant CO2. Now the CO2 level is again dangerously low, as oceans continue to tie it up, while volcanic activity is decreasing due to extremely thick tectonic plates which do not easily allow lava to get through. It produces a fluorescent, blue-green pigment in water. The pigment glows for insects, which see at a higher frequency than humans, causing them to pick up the bacteria and carry them around on their feet.
It produces a fluorescent, blue-green pigment in water. The pigment glows for insects, which see at a higher frequency than humans, causing them to pick up the bacteria and carry them around on their feet. The oldest existing mushroom is the puffball. It would have been something similar about 200 to 300 million years ago. Conifer trees evolved about 300 million years ago. It is quite likely that puffballs evolved at the same time.
The oldest existing mushroom is the puffball. It would have been something similar about 200 to 300 million years ago. Conifer trees evolved about 300 million years ago. It is quite likely that puffballs evolved at the same time.
 This assures a wide and random distribution as a method of preventing predators and diseases from adapting to them.
This assures a wide and random distribution as a method of preventing predators and diseases from adapting to them. A few spores would drop straight down, but the primary form of dissemination, at least originally, was animals eating them. So the spores have an attractive flavor for animals.
A few spores would drop straight down, but the primary form of dissemination, at least originally, was animals eating them. So the spores have an attractive flavor for animals. Grass is usually short enough to allow other species to grow above it including woody plants, flowering plants, mammals and mushrooms. Mushrooms could now use wind which blows through the grass, and they evolved gills for that. Gills allow spores to stay in place until wind sweeps them out and carries them long distances. Flowering plants were so rare during dinosaur years that none had been detected until a few years ago. But they were in place ready to take advantage of the marvelous ecology which grass created.
Grass is usually short enough to allow other species to grow above it including woody plants, flowering plants, mammals and mushrooms. Mushrooms could now use wind which blows through the grass, and they evolved gills for that. Gills allow spores to stay in place until wind sweeps them out and carries them long distances. Flowering plants were so rare during dinosaur years that none had been detected until a few years ago. But they were in place ready to take advantage of the marvelous ecology which grass created. Apparently, not all of the endomycorrhizal mushrooms (those growing within tree roots) began during the dinosaur years. Chanterelles grow within conifer roots, and they have ridges which function like gills with increased exposure to take advantage of the reduced wind in conifer forests. They probably evolved with modern mushrooms, as the exposed spores would have similar requirements. Truffles grow on oak roots, while oaks and other broadleaf trees did not exist during the dinosaur years.
Apparently, not all of the endomycorrhizal mushrooms (those growing within tree roots) began during the dinosaur years. Chanterelles grow within conifer roots, and they have ridges which function like gills with increased exposure to take advantage of the reduced wind in conifer forests. They probably evolved with modern mushrooms, as the exposed spores would have similar requirements. Truffles grow on oak roots, while oaks and other broadleaf trees did not exist during the dinosaur years. Coral mushrooms have spores which are even more exposed than the Chanterelle. Their exposed spores indicate that they would have evolved at the same time as gilled mushrooms, because the microstructure and physiology required for such exposed spores would be the same for gilled mushrooms, chanterelles and corals.
Coral mushrooms have spores which are even more exposed than the Chanterelle. Their exposed spores indicate that they would have evolved at the same time as gilled mushrooms, because the microstructure and physiology required for such exposed spores would be the same for gilled mushrooms, chanterelles and corals. These two boletes would appear to be minor variations in genotype, but they could be phenotypic variants of the same genotype. Boletus edulis appears to produce phenotypes which adapt to conditions. Such ancient evolution appears to produce phenotypic variation which looks like genotypic variation due to a high degree of stability in response to environmental demands. It produces biochemical variations like a chameleon. In shaded areas, the color can be darker. The spores vary from white to brown.
These two boletes would appear to be minor variations in genotype, but they could be phenotypic variants of the same genotype. Boletus edulis appears to produce phenotypes which adapt to conditions. Such ancient evolution appears to produce phenotypic variation which looks like genotypic variation due to a high degree of stability in response to environmental demands. It produces biochemical variations like a chameleon. In shaded areas, the color can be darker. The spores vary from white to brown. Chew marks are often observed on the surface showing that animals walked away after one bite. It is usually squirrels which chew into the top of boletes, while mice eat the spore tissue under the cap. When Boletus edulis gets large, squirrels chew the edge of the cap, where they encounter the flavor of the spore area.
Chew marks are often observed on the surface showing that animals walked away after one bite. It is usually squirrels which chew into the top of boletes, while mice eat the spore tissue under the cap. When Boletus edulis gets large, squirrels chew the edge of the cap, where they encounter the flavor of the spore area.