 |
|
Research Photo Pages
|
Polar flagella would have evolved before terrestrial life began, 541 million years ago, because exotic structures were easy to produce during early evolution. Filamentous fungi show extremely frivolous structures which evolved early in evolution, such as innumerable variations of the clamp connection. Not only was there less competition and related demands early on, but less specialization allowed more varied alternatives. All biological life is differentiating itself out of existence through over-specialization, which is why so many species die out when environmentally stressed. Biological life could end in 10 million years or less due to over-specialization. There is another type of flagella called peritrichous flagella, which surrounds some bacteria. That flagella is designed for moving through viscous liquids, and it is more recent. The difference stems from modern biology creating viscous liquids with nutrients, while in the ancient world water with few nutrients was the primary liquid for biology. Scattered nutrients in an ancient sea gave P.f. a more versatile nutrition and physiology. Nowdays, bacteria specialize more, because they have specific habitats with consistent nutrients. Evidence of P.f. moving out of an ancient sea onto land is in the fact that it is the most common and well adapted soil bacterium. P.f. adapted to soil better than any other bacteria, because it was the first bacterium in the soil. Pseudomonads are linked together in pairs under nonaqueous conditions, which is the basis for the name. This allows one of the bacteria to break open and release enzymes near the other one. Whereas molds produce enzymes on their surface, bacteria have to break open to release enzymes. P.f. breaks down proteins in the soil, which is highly significant where ground freezes. Ice crystals break apart most cells in the soil including some plant roots, fungi and insects. P.f. is the first bacterium to act upon the resulting cell debris in the spring. But it will also grow on extremely diverse nutrients. The isolation medium for it is glycerol and nitrate. Nothing else noticeable grows on that combination. Under nonaqueous conditions, P.f. produces a pink, nonwater-soluble pigment. This pigment shows up when growing P.f. on glucose and agar. Most bacteria would die for a pigment, but pigments are usually too extravagant for them. P.f. has two. Pigments allow insects to pick up the bacteria and carry them around. Also indicative of evolutionary age is symbiosis. The oldest species accommodate their environment the best. P.f. is like a slave for plants feeding them organic nitrogen. For example, when grains such as wheat are planted in the spring, the leaves start to turn yellow due to a shortage of nitrogen, because P.f. hogs up the nitrogen early in the spring. After about a month of growth, the leaves turn dark green, as nitrogen becomes available. P.f. autolyzes (self-destructs) and releases nitrogen to the grain in the nick of time. I find that P.f. also autolyzes (breaks apart) below pH 5.0. This allows other species to feed on it by producing acid. The morel mushroom does this. It excretes a small amount of acid for feeding on P.f. Other mushrooms get most of their nitrogen from bacteria, but it is doubtful that they excrete acid. They probably wait for the bacteria to autolyze for other reasons such as drying. Plant roots produce some acid, which allows them to feed on P.f. Mushrooms can then exploit the nitrogen along with plant roots.
Puffball mycelium grows under the ground for several decades, continuously expanding to spread as much as a hundred meters. I have seen it emerge from the same area, meaning the same mycelium, at an interval of thirty years. Staying under the ground is a method of preventing insects and disease from developing against it.
These varied puffballs all appear to be phenotypes of the same genotype, because they appeared near each other within two months, which means they grew from the same mycelium. It appears that the puffball can change its phenotypes without going through genetic recombination, unlike other species. It's possible that the mycelium sectors as a method of creating phenotypes. This would be like cells changing phenotypes to create tissues but then going beyond the tissue to create individual phenotypes. The puffball produces no visible gene exchange. In general, ancient species do not need much gene exchange, because they acquired very high survivability through versatility. Extreme phenotypes appear to be a major element of the versatility of the puffball. It's lack of gene exchange creates a need for highly varied phenotypes. Puffballs do not appear in the same area for many years, which would be a means of preventing predators or diseases from developing against them. Such extremely ancient organisms produce that sort of versatility. The mechanism is mysterious. My guess is that the spores are presssure sensitive, so that only those that get high in the atmosphere can germinate. This assures a wide and random distribution.
Measuring low pressure would not be based on force but evaporation rate. A low volatility substance would evaporate at a controllable rate at low pressure. Measuring concentrations of metabolites is what cells normally do. Boletus edulis
When B.e. evolved, the only animals around were small and large including shrew-like mammals and dinosaurs. Nonwoody plants produced that result. These plants would have created a very thick brush. Dinosaurs had to be large to walk through that brush. Mammals had to be small to go under it. Mushrooms could not grow in that brush. The surface of the ground under the brush would have had a thick layer of undecayed debris. Puffballs probably grew around the edges of the brush feeding on bacteria associated with plant roots. There were no mountains during the dinosaurs years, because tectonic plates were thin. When plates collided, they would stick together and form low hills instead of mountains. They were all stuck together forming a large mass called Pangaea. Pangaea was starting to break up into continents when the dinosaurs died out 65 million years ago and modern biology began. The nonwoody brush would not grow on hills, because water would run off too easily. This left the hills open allowing conifer trees to evolve on them. Conifers have prickly leaves and acrid chemicals to prevent dinosaurs from eating them. Conifers left space on the ground beneath them for mushrooms to evolve in. Boletus edulis grew into the roots of the conifers establishing a symbiotic relationship to the plants. Wind cannot easily get into conifer forests, so B.e. could not exploit wind very well for spore dissemination. One reason why B.e. did not evolve better spore dissemination is because it must be perennial to grow in tree roots, so it is stuck in a fixed location from year to year. The primitive structures of B.e. indicate that its evolutionary age is about the same as the conifers which it grows on, which goes back 300 million years. It's possible that puffballs evolved on low hills before conifers. This would make the evolutionary age of puffballs more than 300 million years. After conifers evolved, puffballs found space around he edges of the forests and the nonwoody brush. Almost all terrestrial biology changed drastically as the dinosaurs died out. The drastic changes were caused by grass. Grass shoved out most of the nonwoody plants freeing both plants and animals from the oppressive brush that hindered growth for most species. The critical characteristic of grass is that it can absorb water above the ground, while other plants cannot. The long thin leaves of grass collect dew drops which run down the leaves to a point near the ground where water is absorbed. This allows grass to grow on dew and use light rains efficiently.
These two boletes appear to be minor variations in genotype. Very limited spore dissemination and gene exchange may result in Boletus edulis producing localized patches of genotypic difference, much like the morel. But true to its 300 million years of evolution, it apparently produces biochemical variations, like a chameleon. In shaded areas, the color can be darker. The spores vary from white to brown. When spore dissemination is limited, as with the bolete or the morel, small changes in genotype occur in local areas. But when spore dissemination is very good, as with the puffball, minor differences in genotype cannot develop, because one genotype will prevail over the others. Then phenotypic differences get more exaggerated, which appears to be the case with the puffball. Structural tissue is the same, and very unusual, for boletes and puffballs. The tissue has a gummy texture without fibrous material. This means water and nutrients must diffuse from cell to cell, which is a slow process. Therefore, it takes two weeks for the Boletus edulis sporocarp to form, but only one or two days for Agaricus. Bolete tissue is very slow at drying. As a result, boletes often survive for two months, while Agaricus tends to dehydrate and disintegrate in one week.
Both the bolete and the puffball evolved away all flavor from the structural tissue, so animals will not eat it. Chew marks are often observed on the surface showing that animals walked away after one bite. It is usually squirrels which chew into the top of boletes, while mice eat the spore tissue under the cap. When Boletus edulis gets large, squirrels chew the edge of the cap, where they encounter the flavor of the spore area. As Boletus edulis gets large, the bulb on the stem disappears. This is because larger animals are then depended upon, and not a single molecule is wasted on a form that is not needed with such an ancient mushroom. Gilled mushrooms cannot evolve away flavor, because there are too many molecules which would need to change, and evolution progressed to more specialized functions. Therefore, gilled mushrooms have to produce a toxin, if they need to keep animals from eating them. The flavor of Boletus edulis increases as the mushroom dries, and the structural tissue picks up flavor upon drying. This effect probably evolved to enhance the tendency of animals to eat the mushroom as it ages. When Boletus edulis is dried slowly it acquires more flavor than when it is dried rapidly, which indicates that a biochemical process is involved in enhancing the flavor. The Morel
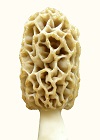
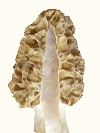
Micromorphology is still yest-like for the morel. The tissue and sclerotial cells are rounded, looking like potatoes. Apparently, the rounded shape leaves space between cells which is occupied by fluids. Morel tissue is unusually crumbly, and this is probably because there is less adhesion between rounded cells than there would be with cells which fit together better with sharp corners.
In other words, the morel has not had enough time to develop usual multicellular characteristics since it changed from a single-celled yeast.
|
 P.f. began in an ancient sea about a billion years ago. The evidence is that it is still the most aquatically adapted bacterium. It has two polar flagella for high speed motion in water. It also produces a fluorescent, blue-green pigment in water. The pigment glows for insects, which see at a higher frequency than humans, causing them to pick up the bacteria and carry them around on their feet.
P.f. began in an ancient sea about a billion years ago. The evidence is that it is still the most aquatically adapted bacterium. It has two polar flagella for high speed motion in water. It also produces a fluorescent, blue-green pigment in water. The pigment glows for insects, which see at a higher frequency than humans, causing them to pick up the bacteria and carry them around on their feet. The oldest existing mushroom is the puffball. It would have been something similar about 300 million years ago. Plants were producing a lot of round aggregates of cells back then and appeared to be evolving into fungi. My impression is that numerous different fungi evolved from plants independently after terrestrial life began. So why do they have similar cell walls if independent? This question would have been unanswerable a couple decades ago, but what scientists have learned from genetic engineering is that horizontal transfer exchanges genes between all species. Viruses, bacteria and fungi serve as the carriers. This is noticeable in wildflowers. They all seem to have a combination of the same basic scents, even when they are not related. Horizontal transfer apparently carried some of the same genes throughout the wildflowers. This means that plants could evolve into a myriad of fungi by picking up the cell wall genes from other fungi. By evolving from plants instead of other fungi, more versatility was possible. Evolution cannot go backward from more specialization to less specialization. Restarting evolution from plants would be like starting from stem cells allowing greater diversity than all fungi evolving from one point.
The oldest existing mushroom is the puffball. It would have been something similar about 300 million years ago. Plants were producing a lot of round aggregates of cells back then and appeared to be evolving into fungi. My impression is that numerous different fungi evolved from plants independently after terrestrial life began. So why do they have similar cell walls if independent? This question would have been unanswerable a couple decades ago, but what scientists have learned from genetic engineering is that horizontal transfer exchanges genes between all species. Viruses, bacteria and fungi serve as the carriers. This is noticeable in wildflowers. They all seem to have a combination of the same basic scents, even when they are not related. Horizontal transfer apparently carried some of the same genes throughout the wildflowers. This means that plants could evolve into a myriad of fungi by picking up the cell wall genes from other fungi. By evolving from plants instead of other fungi, more versatility was possible. Evolution cannot go backward from more specialization to less specialization. Restarting evolution from plants would be like starting from stem cells allowing greater diversity than all fungi evolving from one point.
 Perhaps other airborne mushroom spores require atmospheric conditions, of which low pressure is the only apparent mechanism, for evading diseases. Agaricus avoids growing in the same area from year to year. What keeps the spores from germinating in the near vicinity of the mushroom which produces them? It might be atmospheric conditions being required for spore germination. Coprinus comatus, however, liquefies to drop some of its spores in the same area, which means its spores do not need to be exposed to atmospheric conditions.
Perhaps other airborne mushroom spores require atmospheric conditions, of which low pressure is the only apparent mechanism, for evading diseases. Agaricus avoids growing in the same area from year to year. What keeps the spores from germinating in the near vicinity of the mushroom which produces them? It might be atmospheric conditions being required for spore germination. Coprinus comatus, however, liquefies to drop some of its spores in the same area, which means its spores do not need to be exposed to atmospheric conditions.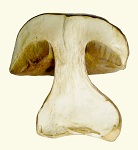 Also ancient is the mushroom Boletus edulis (B.e.), which is highly valued for food. B.e. does not have gills. Instead it has tubes under the cap with spores in them. A few spores would drop straight down, but the primary form of dissemination was animals eating them. So the spores have an attractive flavor for animals.
Also ancient is the mushroom Boletus edulis (B.e.), which is highly valued for food. B.e. does not have gills. Instead it has tubes under the cap with spores in them. A few spores would drop straight down, but the primary form of dissemination was animals eating them. So the spores have an attractive flavor for animals. Grass is usually short enough to allow other species to grow above it including woody plants, flowering plants, mammals and mushrooms. Mushrooms could now use wind which blows through the grass, and they evolved gills for that. Gills allow spores to stay in place until wind sweeps them out and carries them long distances. Flowering plants were so rare during dinosaur years that none had been detected until a few
Grass is usually short enough to allow other species to grow above it including woody plants, flowering plants, mammals and mushrooms. Mushrooms could now use wind which blows through the grass, and they evolved gills for that. Gills allow spores to stay in place until wind sweeps them out and carries them long distances. Flowering plants were so rare during dinosaur years that none had been detected until a few  Apparently, not all of the endomycorrhizal mushrooms (those growing within tree roots) began during the dinosaur years. Chanterelles grow within conifer roots, and they have ridges which function like gills with increased exposure to take advantage of the reduced wind in conifer forests. They probably evolved with modern mushrooms, as the exposed spores would have similar requirements. Truffles grow on oak roots, while oaks and other broadleaf trees did not exist during the dinosaur years.
Apparently, not all of the endomycorrhizal mushrooms (those growing within tree roots) began during the dinosaur years. Chanterelles grow within conifer roots, and they have ridges which function like gills with increased exposure to take advantage of the reduced wind in conifer forests. They probably evolved with modern mushrooms, as the exposed spores would have similar requirements. Truffles grow on oak roots, while oaks and other broadleaf trees did not exist during the dinosaur years. Coral mushrooms have
Coral mushrooms have 



 The significance of morphological evolution is demonstrated with the morel. It shows that macromorphology changes much more easily than micromorphology or physiology. It's macromorphology is crude and loosely controlled, often consisting of cells hanging off the side. There has been no change in microstructure from the yeast ancestor. The spores would be exactly the same in the yeast ancestor. The spore surface pattern was said to be the same for the morel and two cup fungi (Dsicinia and Disciota, Nancy Smith Weber), which means those cup fungi would be morel relatives from earlier ice age cycles. The morel re-evolves from a yeast during each
The significance of morphological evolution is demonstrated with the morel. It shows that macromorphology changes much more easily than micromorphology or physiology. It's macromorphology is crude and loosely controlled, often consisting of cells hanging off the side. There has been no change in microstructure from the yeast ancestor. The spores would be exactly the same in the yeast ancestor. The spore surface pattern was said to be the same for the morel and two cup fungi (Dsicinia and Disciota, Nancy Smith Weber), which means those cup fungi would be morel relatives from earlier ice age cycles. The morel re-evolves from a yeast during each  The morel does not have sufficiently developed signal molecules for precise control of cell positions. Evidence of the lack of feedback information is in its inability to respond to gravity and grow vertically. Whichever direction a morel starts to grow is the direction in which it keeps growing. The morel can't measure gravity until it acquires sufficient signal molecules for feedback information. Almost everything else which emerges from the soil measures gravity for vertical growth.
The morel does not have sufficiently developed signal molecules for precise control of cell positions. Evidence of the lack of feedback information is in its inability to respond to gravity and grow vertically. Whichever direction a morel starts to grow is the direction in which it keeps growing. The morel can't measure gravity until it acquires sufficient signal molecules for feedback information. Almost everything else which emerges from the soil measures gravity for vertical growth.