|
|
Research Photo Pages
|
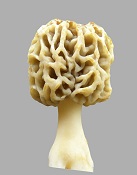
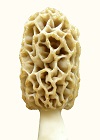
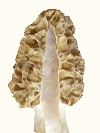
The pairs below are genotypically the same but phenotypically different. They grow near each other, which means they exchange genes and become genetically the same. Morels
Bindweed
Spurge
Echinacea
Liatris
Phenotypic variation as an adaptation mechanism is an important biological phenomenon which was not previously known to exist. When aware of it, it is easy to see in plants. It would exist in animals also, such as muscle cell variation. It would be quite extreme in yeasts, which is where the morel acquired it. It is observed as variations in a population which would be genetically homogeneous. Populations become homogeneous over time due to gene exchange. Variations must be introduced from outside sources. The source of differences is diverse environmental conditions. In other words, inbreeding causes offspring to become genetically similar. Inbreeding means lack of outside diversity. When variations are introduced into a population, they spread according to the mendelian pattern, which means from parents to offspring with half of the genes from each parent. Gradually, the variations blend uniformly into the population. By contrast, phenotypic variation is observed as randomized distribution of variations, and it does not disappear over time by blending into the population. The test of phenotypic variation is randomization of variations in place of the usual Mendelian variations following lines of inheritance. A random distribution of variations is observed. Another test of phenotypic variation is variation being sustained in a population without blending in through gene exchange. Biological variations are created by differences in environments. An example would be one type of alligator adapting to a swamp which has a lot of grass, and it eats small prey. A different type of alligator adapts to a large open bay, and it eats large prey. So the two types evolve slight differences in size and so forth. Those differences are valuable for survival under diverse conditions, so they are remixed through gene exchange, when the different types come together. Nature invests heavily in gene exchange (as sexuality) so that differences can be remixed into new combinations for improved adaptability. The flowers on plants have that purpose. Flowers attract insects which carry pollen from plant to plant, so differences can be mixed into new combinations. Animals also invest heavily in sexuality producing dramatic colors and courtship rituals to promote gene exchange, so differences can be promoted for better survival under a diversity of conditions. But gene exchange does not create differences, it can only remix what already exists. The mixing process homogenizes the gene pool. Over time, all offspring become similar, unless new sources of variation are added, and they must come from a diversity of environments. A qualifier must be added here. The remixing of genes during gene exchange does produce individual combinations in new ways, but pre-existing genes must be used. The evolution of new genes is a slow process that occurs in response to variations in environmental conditions. How the mixing occurs is evident in human ethnic groups. A few centuries ago, humans were quite isolated and did not move around much. So ethnic groups acquired identifiable characteristics. When they started to travel to different areas and mix with other persons, the differences started to disappear. Notice that an ethnic group acquires its characteristics because gene exchange homogenizes the group, when external sources of variation are limited. In other words, gene exchange does not create diversity, it destroys diversity. Diversity is generated by varied environmental conditions. A lot of scientists assume otherwise. They assume diversity is a product of gene exchange. An alternative to the process of using gene exchange to remix characteristics is phenotypic variation. It is a mechanism used by yeasts because of their limited gene exchange. Yeasts are fungi which adapted to sugary solutions during the era of modern plants beginning about 65 million years ago. In sugary solutions, yeasts lost the ability to disseminate easily. External spores were converted to internal spores, so single cells could form spores. Then the wind could no longer carry spores from one group of cells to another. The limited gene exchange left yeast with reduced ability to adapt to variations in environmental conditions. So to survive under a variety of conditions which they might encounter, they used phenotypic variations. So what is phenotypic variation? It is a difference in appearance or function for types which are genetically the same. For example, the morel forms thick-tissued and thin-tissued mushrooms to cope with variations in weather. Another example of phenotypic variation is embryonic development. Different types of cells are created, even while they are all genetically the same. A muscle cell is genetically the same as a fat cell. The differences are phenotypic, not genotypic. The way it occurs is by repressing some genes and using others. Each cell type uses a different combination of genes, even though all of the cells have the same reservoir of genes. The morel mushroom evolved from a yeast and carried phenotypic variation from its ancestor. It had to continue to use phenotypic variation, because its ascospores limit gene exchange.
The morel spores are formed inside of cells (asci) on the surface of the mushroom. A force propels the spores out of the asci. The force is created by shrinkage of tissue upon drying. Morel spores are very heavy, which allows them to function better as projectiles. They often contain more than twenty nuclei, which increases their mass. Some spores stay on the surface of morels to be picked up by wind. But wind is not used effectively by morels. The surface of morels lacks the aerodynamic advantages of gills, which allow wind to sweep out a large number of spores; and the morel environment is surrounded by trees and brush which reduce wind and block spore movement. Another problem with dissemination of spores by morels is that there are large amounts of space between localized populations of morels, because the soil must be sandy or similar, which is usually limited to river basins. There is a lot of diversity of morel types, and those differences are readily observed over distances of a few hundred miles. But spores are not easily carried by wind over those distances. Regional differences are sustained for this reason. Otherwise, there would be more homogenization of types, as observed with other mushrooms. So the morel had to continue to use and develop phenotypic variation as an adaptation mechanism. It did that by producing a large number of variants which have the same genotype. Some of those variants are better adapted to a particular set of environmental conditions than others. Here's a subtle point which is the key to understanding this subject. If the morel can produce so many variants phenotypically, why not do it genotypically? After all, there has to be a gene for each phenotype. There are two answers to that question. One is time. Speed is the precious commodity of gene exchange. If genes can be rapidly rearranged into new combinations, then changing environmental conditions can be met with new functions. But if diverse options cannot be rapidly brought together and remixed, then a large number of options must be sustained continuously, which is phenotypic variation. The other answer is that genotypic adaptation involves loss of genes. All of the disadvantageous genes are discarded. But gene exchange allows a complete variety of genes to be reacquired from their original sources of diversity. If however, gene exchange is too limited, genes which are lost cannot be reacquired easily, and then the whole diversity of options must be carried through each generation, as occurs with phenotypic variation. The cumulative effect of gene exchange is adaptation by improved function. The cumulative effect of phenotypic variation is adaptation by multiple options. Improved function is more adequate and complex than multiple options. So the phenotypic variation of Morchella is a disadvantageous necessity resulting from the use of ascospores and a carry-over from its yeast ancestry. Yeast Fermentation A place where this information is most important is yeast fermentation. With fermentation, the amount of alcohol and acetic acid and their ratios is always important. Yeast vary in such quantities based on intrinsic characteristics, because varied environments require different responses. But yeast physiologists can never quite pin down the precise characteristics of their cultures, and they never knew why. The reason is phenotypic variation, which they were unaware of. Knowing this, yeast can be prevented from changing their phenotypic characteristics by preventing them from forming spores, which is not difficult. Once the phenotypes are stabilized, they can be selected for desired characteristics. Epigenetics is complex phenotypic variation In its simplest form, as shown in these photos, phenotypic variation is determined when an individual forms through reproduction. A more complex version is visible with the puffball, where at least four phenotypes spring from the same mycelium in sequence separated by several weeks. Even more complex is phenotypic change which is carried through more than one generation, and it is called epigenetic inheritance.
This type of puffball would be the central line of evolution. Numerous variants branch off from the central line. The central line of the puffball had similar characteristics going back 300 million years with refinements occurring over time. With yeasts and the morel, the phenotype is presumably determined during karyogamy. With the puffball, biochemical signals change phenotypes during stages of growth. What is being shown with epigenetic studies is that the signals can be induced through nutrition to create phenotypes that carry through more than one generation. See Morel Phenotypte Photo Page.
|