|
|
Research Photo Pages
|
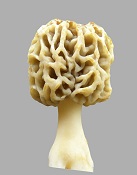
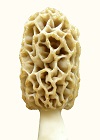
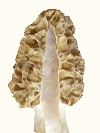
The morel mushroom is highly varied in its morphology, both genotypically and phenotypically. Morels are quite isolated over short distances because of their difficulty in getting spores disseminated. The spores must be propelled out of the ascus with a force created through shrinkage of tissue upon drying. The propelled spores travel up to a meter of distance only. Very seldom does wind pick up the spores; and then the spores have very little chance of ending up in another patch of morels to create genetic interactions. So the genes do not easily interact over some distance. Within a patch of morels, gene exchange does not occur between mycelia of different phenotypes. A wall develops between mycelia of different phenotypes to prevent phenotypic differences from being destroyed. The wall between different mycelial types has been referred to as a meld in early research, not knowing why it occurs. However, the "meld" does not develop about 2% of the time, which allows some gene exchange to occur without disrupting most phenotypic differences. In the upper plains of the U.S., the harsh conditions result in observable differences in morel genotypic morphology about every 100 miles of separation or less. Under the more ideal conditions of the west coast, morphological differences are much less noticeable. Then within each patch, phenotypic differences in morphology are extreme under harsh conditions but less so under more favorable conditions. The genetic differences are observable through the phenotypic differences as pattern trends. These two morels are from the same patch, which means they are genotypically the same but phenotypically different:
The morel emerged from the ground about 20 thousand years ago, since it re-evolves at the base of trees during each ice age. Ice ages occur like clockwork every 100 thousand years. Cold, humid air sweeping off the ice sheet must keep the mycelium of a filamentous yeast from dehydrating. That process cannot occur apart from the ice sheet, because the leaves of the trees must get sunshine, while the base of the trunk is continuously hydrated by cold, damp fog. The extreme morphological differences are needed to cope with the difficulty of getting spores out. The tissue must stay hydrated for about three days while spores form; and then drying must occur to cause the tissue to shrink and create a force that propels the spores out. So the morphological differences include fast drying and slow drying types plus various shapes for coping with surrounding environments. The morels are often embedded in vegetation including grass and stemy brush. While the morphological evolution of morels is rapid and extreme, the physiology is highly immune to variation. Physiology cannot easily evolve, because the inside of cells requires precise location of enzymes which are attached to membranes. It's like thousands of assembly lines packed together. The reactants must not diffuse away but must move the shortest possible distance from one enzyme to another. So the morel has a lot of disadvantageous physiology including residual autolysis, which almost all bacteria and yeast use to recycle nutrients. The autolysis results in one side of the morel deteriorating as it ages and picking up gram negative bacteria (which prefer high-nitrogen nutrients). The gram negative bacteria will make people sick, if they pick old morels, because of endotoxin which does not degrade with cooking. The use of acetic acid to kill bacteria and feed upon them requires marginal conditions, were the mycelium is spread thinly over much area. When the mycelium is concentrated under artificial conditions, the accumulation of acid overwhelms the mycelium. The ease by which morel morphology evolves indicates that changes in shape simply require counting the number of cells in each direction, which is a lot easier to do than changing physiology. What then is counting cells in each direction? To control morphology, each cell needs to have its own identity and needs to know what cells surround it, while they have a different identity. What then is cell identity? A clue is provided by injured tissue of humans. With a simple cut, cells can grow back with no evidence later. But if the tissue is significantly damaged, a scar develops. The scar says the cells cannot determine their required surrounding environment and must form a generic tissue. The scar tissue says each cell has an identity and a need to know the identity of the surrounding cells. That means signaling between cells is required. The signals would need to be RNA to contain the required amount of information. So there needs to be a lot of DNA devoted to producing the signal RNA. Where in a genome is all of that information? A very large part of any genome has no identifiable purpose. That means there is a lot of DNA available for producing the identity RNA. Transposons are sections of DNA that move around a lot. They do not have identifiable functions. They are said to be "selfish genetic entities" in that they do not seem to contribute to cell functionality. Why would they not be eliminated through evolution? Probably because they have an unidentified purpose. That purpose of the transposons seems circumstantially to be to create the RNA for cell identity. If so, the reason why the transposons move around so much on the DNA would be to promote evolution of morphology. Evolution creates hot spots for change and cold spots which prevent change. There are genes locked in place which do not change over millions of years. They are normally surrounded by proteins; and if they mutate, the changed base subunit will be excised out and replaced with the proper base which matches the corresponding base across from it. An example of how the process works was when amphibians crawled out of the oceans and evolved bones needed to overcome gravity for land motion. Then some of the amphibians moved back into the oceans and evolved into boney fish. The fish that stayed in the oceans never evolved bones but used cartilage for structural purpose. Then the boney fish moved back out of the oceans and evolved into mammals. The mammals could evolve more complexity than the amphibians that never entered the oceans. Those amphibians evolved into dinosaurs and reptiles. The point is that moving into the oceans and back out again allowed mammals to evolve in extreme ways; so they could produce more complexities including live birth replacing eggs. That's because moving into the oceans and back out again required so much rapid and extreme evolution that it stripped the proteins and related protections off a large part of the DNA, where rapid evolution was needed. The reptiles had so much of their DNA protected from mutations that they could not easily adapt and their evolution stay very primitive. Which explains why transposons are so active. They promote a lot of rapid evolution of morphology. Transposons serving the function of identity control explains why they do not disappear in evolution. The assumption that they can be sustained selfishly without contributing to evolution is not logical, since all DNA that does not contribute to evolution is degraded over time, at least if it is active and in an area that is not highly protected. And the absence of an apparent function for transposons indicates that creating identity RNA is the purpose. Creating RNA as the only function would look like the absence of a function.
|